
Содержание Перестройка бронхиального эпителия при хроническом бронхите.Определение непрямым методом Кунса иммуноглобулинов классов A, G, М показало их определенную динамику и изменение взаимоотношений между ними. Для первой формы, где взаимоотношения между клетками бронхиального эпителия относительно сохранены и имеются лишь признаки гиперсекреции, в инфильтрате преобладают лимфоциты, лазмоциты и гистиоциты. В плазматических клетках выявляется Ig A. Секреторный иммуноглобулин А локализуется в апикальных отделах реснитчатых клеток. Клетки, синтезирующие Ig M и Ig G, встречаются редко. Вторая форма характеризуется нарастанием альтеративных изменений в бронхиальных клетках, изменением соотношения между ними в пользу гиперплазированных и гипертрофированных бокаловидных клеток, при интактных пока базальных клетках, сопровождается изменениями местного иммунитета. В инфильтрате, помимо лимфоцитов и плазматических клеток, появляются эозинофилы, тучные клетки, по-линуклеарные нейтрофилы. Число глобулинопродуцирующих клеток, с преобладанием Ig А, значительно увеличивается. В капиллярах активируются эндотелиоциты, которые выглядят гипертрофированными, в них появляются пиноцитозные пузырьки, цитоплазматические выросты. В процесс вовлекается железистый аппарат БВБ, где также отмечается гиперплазия и гипертрофия клеток с накоплением ШИК-положительной слизи в концевых отделах и выводных протоках, лимфоплазмоклеточная инфильтрация стромы желез. В слизистых клетках желез и в апикальной зоне реснитчатых клеток выявляется секреторный иммуноглобулин А. В дальнейшем, при третьей форме, процесс гиперсекреции бокаловидных клеток переходит в их дистрофию. Практически полностью нарушается реснитчатый покров. Частично реснитчатые и бокаловидные клетки замещаются эпителиоцитами полигональной формы. В эпителии редки клетки, содержащие Ig А, в собственной пластинке слизистой оболочки доминируют плазмоциты, маркированные Ig G. В стенках мелких вен выявляются иммунные комплексы, Ig G и М. Авторы трактуют эти изменения как смену «первой линии защиты» — второй, где ситуацию контролирует Ig G, заменяющий Ig А. Способность Ig G комплексироваться с антигеном с активацией комплемента по классическому пути создает предпосылки для развития иммунокомплексной реакции. В этих условиях нарушение микроокружения базальных клеток, фиксируемое авторами, с изменением дифференцировки и регенерации покровного эпителия, является «существенным звеном в самоподдержании воспалительного процесса». При четвертой форме у больных находят метаплазию эпителия бронхов в многослойный плоский, инфильтрат собственной пластинки слизистой оболочки — скудный, основу его составляют фибробласты, плазматические клетки и лимфоциты. Собственная пластинка слизистой оболочки бронхов склерозирована. Имеется периваскулярный склероз. Единичные плазматические клетки маркированы Ig А, превалируют клетки с Ig G в цитоплазме, в бронхиальном эпителии секреторный иммуноглобулин А отсутствует.
Таким образом, перестройка бронхиального эпителия при хроническом бронхите сопровождается утратой способности синтезировать секреторный иммуноглобулин А. Это снижает свойства слизистой оболочки (БВБ), тем самым создается порочный круг, в котором важную роль играет состояние местного иммунитета и иммуннопатологические реакции. Развитие работ этого направления внесло некоторые уточнения в характер распределения иммуноглобулинов, в целом подтвердив, что недостаточность местных гуморальных факторов связана с нарушением взаимоотношений в эпителиальном компоненте БВБ. Не вступая в дискуссию по поводу некоторой условности деления описанного процесса на формы, следует отметить, на наш взгляд, важный факт системной перестройки бронховаскулярного барьера, всех его компонентов при развитии хронического воспаления. Количественные и качественные изменения в представительстве иммунокомпетентных клеток, заселяющих стромальный компонент БВБ, являются отражением общих системных нарушений иммунитета, предваряющих и сопровождающих развитие и течение хронического бронхита. Убедительно показано наличие дисфункции центральных звеньев иммунитета, наличие тимической недостаточности со снижением количества Т лимфоцитов и их пролиферативной активности, обнаруживаемой тестами с фитогемаглютинином и конканавалином А, особенно при наличии обструктивного синдрома. Недостаточность неспецифических механизмов, в частности фагоцитарного, определяется исходно сниженной функциональной активностью про- и моноцитов костного мозга у этих больных. Снижены фагоцитарная активность и индекс миграции лейкоцитов на фоне угнетенного энергетического метаболизма. Изменения затрагивают и трахеобронхиальные лимфатические узлы. Тяжелое течение хронического бронхита сопровождается меньшим содержанием плазматических клеток, маркированных иммуноглобулинами классов А и М. Наши результаты по подсчету различных типов клеток-эффекторов иммунной системы, локализованных в БВБ на уровне дольки при хроническом бронхите без клинических признаков обострения показали, что инфильтрат формируется не только из лимфоцитов. Суммарно — до 15-20% от общего числа клеток приходится на плазматические, тучные клетки, нейтрофильные и эозинофильные лейкоциты. Изменен не только качественный состав хронической воспалительной реакции, меняется характер взаимоотношений, между клетками — эффекторами. Анализ межклеточных корреляционных связей показал, что при хроническом воспалении бронхов утрачиваются одни и появляются новые варианты кооперативных отношений между клетками иммунной системы. В макрофагах и нейтрофилах высока активность ферментов пентозного шунта и снижена — ферментов гликолиза. В лимфоцитах регистрируется низкая активность одного из ключевых ферментов цикла Кребса — СДГ, более высокая, чем у здоровых людей, активность лизосомального фермента — кислой фосфатазы. Реактивность организма и ферментативная активность иммунокомпетентных клеток коррелируют, по некоторым данным, с формой и степенью дыхательной недостаточности. Согласно иммунологическим тестам возможны три варианта течения хронического бронхита: с нормальной иммунологической реактивностью — катаральная форма; с селективным увеличением числа циркулирующих «ноль» лимфоцитов в сочетании с недостаточностью антител классов А и G — обструктивная форма; с Т иммунодефицитом, увеличением продукции «ноль» лимфоцитов и антител класса М при недостаточности иммуноглобулина А — длительно текущая обструктивная и гнойно-обструктивная форма. — Также рекомендуем «Снижение количества реснитчатых клеток при хроническом бронхите.» Оглавление темы «Барьеры легких при воспалении.»: 1. Генетические факторы инфекционного воспаления. 2. Причины и факторы дефицита в системе иммуноглобулинов. 3. Генетические аспекты специфического воспаления. 4. Морфология легких человека при воспалении. 5. Компоненты барьеров легких при хроническом бронхите. 6. Механизм контроля состава слизи при хроническом бронхите. 7. Перестройка бронхиального эпителия при хроническом бронхите. 8. Снижение количества реснитчатых клеток при хроническом бронхите. 9. Изменения в паренхиме легких при хроническом бронхите. 10. Компоненты барьеров легких при острой пневмонии. |
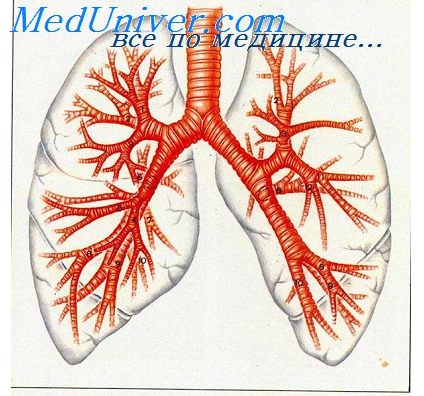
оль эпителия бронхов при воспалении.
Роль эпителия бронхов при воспалении.Этот фрагмент не претендует на полноту, поскольку некоторые вопросы, рассматриваемые в других разделах сайта, в частности, роль лимфоцитов в воспалении, описывается в статьях, посвященных роли иммунной системы. Эпителий бронхов. Большую роль в развитии воспаления бронхов играет повреждение эпителия. Функции его многообразны: во-первых, в эпителии находятся наиболее эффективные антиген-презентирующие дендритные клетки, которые после поглощения антигена мигрируют в региональные лимфоузлы, где контактируют со специфическими Т-лимфоцитами; во-вторых, клетки эпителия экспрессируют разнообразные рецепторы, благодаря которым с эпителиоцитами связываются различные БАВ, клетки воспаления, бактерии и; в-третьих, эпителиоциты активно продуцируют медиаторы, в частности, провоспалительные цитокины, поддерживающие хроническое воспаление. Велика роль эпителия в регуляции экссудации плазмы в просвет бронхов: клетки эпителия, как и другие типы клеток, продуцируют вазоактивные медиаторы экссудации, включая различные лейкотриены (ЛТ), прежде всего ЛТД4, и фактор активации тромбоцитов (ФАТ), причем, первоначальная экссудация имеет защитное действие, поскольку плазма образует гель, который защищает обнаженную мембрану, поэтому имеется связь между степенью повреждения эпителия (аллергеном, поллютантом, бактерией) и выраженностью плазменной экссудации.
Эпителий продуцирует много провоспалительных цитокинов благодаря выраженной метаболической активности, в частности, метаболит арахидоновой кислоты 15-НЕТЕ (15-гидроксиейкозатетраеновая кислота), который стимулирует продукцию слизи и активирует 5-липоксигеназный путь обмена в тучных клетках, простагландины Е, и F2a, GM-CSP (гранулоцитарно-макрофагаяьный колониестимулируюший фактор), активирующий альвеолярные макрофаги, гранулоциты и базофилы, GCSP (гранулоцитарный колониестимулирующий фактор), интерлейкины (ИЛ), в частности ИЛ-1, ИЛ-6 и ИЛ-8 — хемоаттрактант и активатор эозинофилов и нейтрофилов. Одним из главных хемоаттрактантов, выделяемых эпителием со специфическим привлечением моноцитов — МСР-1 (моноцитарный хемоаттрактантный пептид-1). Эпителий бронхов выделяет также другие медиаторы воспаления: эндотелии-1,- активный вазо- и бронхоконстриктор и активатор фибробластов, цитокины группы RANTES из подсемейства полипептидов В или «С-С» (Regulated on Activation, Normal T-cell Expressed and Secreted), привлекающие эозинофилы, различные факторы роста (PDGE — пластиночный фактор роста, BFGF — основной фактор роста фибробластов, IGE — инсулиноподобный фактор роста),- все из них способны принимать участие в ремоделировании дыхательных путей. В последнее время большое внимание уделяется выделению эпителием бронхов окиси азота (N0), которая обладает вазо- и бронходилатирующим действием и ее концентрация по-разному изменяется при различных легочных болезнях. Важным в генезе воспаления является экспрессия различных молекул, прежде всего, молекул адгезии ICAM-I (intercellular adhesion molecnle-I), необходимая для транспорта нейтрофилов и эозинофилов через слизистую в просвет дыхательных путей, а также рецепторов автономной нервной системы, что указывает на влияние автономной нервной системы на функции эпителия и формирование нейрогенного воспаления. Помимо адренорецепторов, мускариновых холинорецепторов (в основном, М3), в последнее время интенсивно изучаются VIP-рецепторы (рецепторы вазоинтестинального пептида), возбудитель которых — VIP -сходен по действию с агонистами и дефицит которого может играть важную роль в патогенезе БА; тахикининовые рецепторы, которые с высокой плотностью находятся на эпителии и опосредуют действие мощных провоспалительных медиаторов — вещества П и нейрокинина А; а также большое количество медиаторных рецепторов, посредством которых различные БАВ модулируют метаболическую активность эпителиоцитов, стимулируют экспрессию других видов рецепторов или, наоборот, тормозят ее. Таким образом, эпителиоциты действуют как трансдукторы между различными сигналами, возникающими в просвете бронха и на поверхности эпителия, и воспалительными событиями в стенке бронха, причем в нормальных условиях эпителий поддерживает баланс между про- и противовоспалительными эффектами. При нарушении эпителиального покрова этот баланс нарушается в пользу преобладания провоспалительных эффектов, в просвет бронхов выходят клетки воспаления и жидкая часть плазмы, и при определенных условиях процесс может принять хроническое персистирующее течение. — Также рекомендуем «Роль эндотелия легочных сосудов при воспалении.» Оглавление темы «Воспаление бронхов.»: 1. Аспирин и циклоспорин при воспалении легких. 2. Механизмы воспаления бронхов и легких. 3. Роль эпителия бронхов при воспалении. 4. Роль эндотелия легочных сосудов при воспалении. 5. Альвеолярные макрофаги. Роль альвеолярных макрофагов в воспалении легких. 6. Эозинофилы. Тромбоциты. Роль эозинофилов, тромбоцитов в воспалении бронхов. 7. Тучные клетки и гистамин. Роль гистамина в воспалении бронхов. 8. Эйкозаноиды. Воспаление бронхов и выделение эйкозаноидов. 9. Серотонин. Функции серотонина при воспалении бронхов. 10. Механизмы инфекционного воспаления бронхов. |
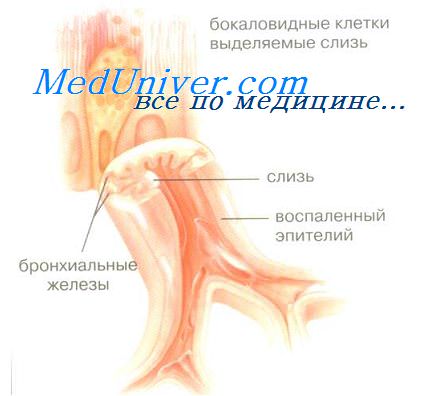
пителиальные клетки и воспаление легких.
Эпителиальные клетки и воспаление легких.Рассматривая роль эпителия дыхательного тракта в системе защиты организма, необходимо выделить следующие его функции: барьерную, эффективный мукоцилиарный клиренс, модуляцию эпителия под влиянием секреторных продуктов как самих эпителиальных клеток, так и других типов клеток, участвующих в воспалении. Эффективное сочетание всех этих функций обеспечивает нормальное функционирование эпителиального слоя легких. Легочный эпителий является морфологическим и функциональным барьером. Гистологически в альвеолярном эпителии определяется наличие комплексов, состоящих из клеточных молекул адгезии и многочисленных специфических белков. Наличие таких комплексов обеспечивает непроницаемость эпителиального барьера для макромолекул и инфекционных агентов, и существенно ограничивает диффузию ионов. Эпителий дыхательных путей является эффективным барьером, препятствующим микробной инвазии, основным этапом которой является контакт микроба с эпителием. Очень мало известно микроорганизмов, способных к адгезии,к нормальному эпителию бронхов и легких. Представителями таких микроорганизмов являются Mycoplasma pneumoniae и Bordetella pertussis. Однако поражение эпителия в результате его механического повреждения (например, при эндотрахеальной интубации) или вирусного инфицирования делает доступным этот процесс для большинства бактерий.
Функция мерцательного эпителия находится в тесной зависимости от различных гуморальных факторов, в том числе клеточного происхождения. Основные из них перечислены в таблице. Кроме того, нижние отделы респираторного тракта содержат многочисленные факторы, продуцируемые эпителиальными клетками. Большая часть этих факторов обладает прямым антибактериальным действием. Ключевая роль эпителиальных клеток в развитии воспаления бронхов и легких определяется их тесным взаимодействием с другими клетками воспаления. Клетки дыхательного эпителия в ответ на стимуляцию обладают способностью продуцировать хемоаттрактанты: IL-8 и RANTES. Эти хемотаксические факторы активируют нейтрофилы, моноциты (или макрофаги), эозинофилы и лимфоциты. Факторы роста (G-CSF; GM-CSF), продуцируемые эпителиальными клетками, могут также индуцировать дифференцировку моноцитов и тучных клеток. Кроме того, эпителиальные клетки участвуют в подавлении воспалительных клеток. Эпителиальные клетки, так же как макрофаги, способны продуцировать TGF-p, который угнетает IL-2-зависимую пролиферацию Т-клеток и продукцию макрофагами других цитокинов. Противовоспалительным эффектом обладают также PGE2 и IL-6, продуцируемые эпителиальными клетками. Выявлено наличие иммуносупрессивной активности альвеолярного эпителия по отношению к Т-лимфоцитам. Эпителиальные клетки продуцируют белок с мол.весом 70кЕ)а, который блокирует пролиферацию Т-лимфоцитов между G1 и S фазами клеточного цикла. Сурфактант также модулирует активность клеток воспаления: подавляет функцию лимфоцитов (цитотоксичность, пролиферацию, синтез иммуноглобулинов) и повышает активность макрофагов. Антилимфоцитарная активность сурфактанта ассоциирована с его липиднои фракцией, в то время как стимуляция макрофагов — с SP-A, который повышает миграционную активность макрофагов, фагоцитоз, внутриклеточный киллинг Staphylococcus aureus и опухолевых клеток. Эпителиальные клетки также способны модулировать функцию гладкомышечных клеток дыхательных путей. Например, PGE2 приводит к бронходилятации, а нейтральная эндопептидаза (NEP) — к бронхоконстрикции. Эпителиальные клетки также влияют на функцию эндотелиальных клеток и выраженность отека дыхательных путей при помощи различных факторов: фермента, разрушающего гистамин (N-метилтрансфераза), 15-липоксигеназы, N0. Эпителиальные клетки могут модулировать рост фибробластов, их пролиферацию. Фибронектин и TGF-(3, секретируемые эпителиальными клетками, непосредственно действуют на миграцию фибробластов, a TGF-(3 и PGE2 модулируют секрецию фибробластами матричных белков. На этот процесс оказывают также влияние и другие факторы, экспрессированные эпителиальными клетками: инсулиноподобный фактор роста (IGF-1), IL-1 и IL-6. Легочный эпителий, без сомнения, участвует в развитии большинства заболеваний дыхательной системы. Повреждение альвеолоцитов I типа — обязательная характеристика многих острых и хронических заболеваний легких, а повреждение дыхательного эпителия — обязательный признак инфекционных процессов, токсических поражений, иммунопатологических процессов и различных хронических заболеваний (БА и хронический бронхит). Повреждение эпителиальных клеток повышает предрасположенность к развитию вторичной инфекции дыхательных путей. — Вернуться в оглавление раздела «Пульмонология.» Оглавление темы «Клетки крови и воспаление легких.»: 1. Механизмы активации эозинофилов. 2. Интерстициальные заболевания легких. Эозинофильная пневмония. Васкулиты. Эозинофильные плевриты. 3. Бронхиальная астма и эозинофилы. Бронхогенные опухоли и эозинофилы. 4. Тучные клетки и базофилы при воспалении легких. 5. Аллергические заболевания, легочный фиброз, опухоли. Макрофаги. 6. Альвеолярные макрофаги. Значение альвеолярных макрофагов при воспалении в легких. 7. Цитокины и биологически активные вещества макрофагов. 8. Второстепенные защитные функции макрофагов. 9. Дендритные клетки и клетки Лангерганса при воспалении легких. 10. Эпителиальные клетки и воспаление легких. |
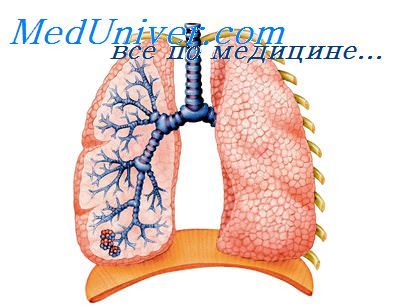
Бронхи
Гистологическое строение бронхов
Снаружи трахея и крупные бронхи покрыты рыхлым соединительнотканным футляром — адвентицией. Наружная оболочка (адвентиция) состоит из рыхлой соединительной гкани, содержащей в крупных бронхах жировые клетки. В ней проходят кровеносные лимфатические сосуды и нервы. Адвентиция нечетко отграничена от перибронхиальной соединительной ткани и вместе с последней обеспечивает возможность некоторого смещения бронхов по отношению к окружающим частям легких.
Далее по направлению внутрь идут фиброзно-хрящевой и частично мышечный слои, подслизистый слой и слизистая оболочка. В фиброзном слое кроме хрящевых полуколец имеется сеть эластических волокон. Фиброзно-хрящевая оболочка трахеи при помощи рыхлой соединительной ткани соединяется с соседними органами.
Передняя и боковые стенки трахеи и крупных бронхов образованы хрящами и расположенными между ними кольцевидными связками. Хрящевой скелет главных бронхов состоит из полуколец гиалинового хряща, которые по мере уменьшения диаметра бронхов уменьшаются в размерах и приобретают характер эластического хряща. Таким образом, из гиалинового хряща состоят только крупные и средние бронхи. Хрящи занимают 2/3 окружности, мембранозная часть — 1/3. Они образуют фиброзно-хрящевой остов, который обеспечивает сохранение просвета трахеи и бронхов.
Мышечные пучки сосредоточены в мембранозной части трахеи и главных бронхов. Различают поверхностный, или наружный, слой, состоящий из редких продольных волокон, и глубокий, или внутренний, представляющий собой сплошную тонкую оболочку, сформированную поперечными волокнами. Мышечные волокна располагаются не только между концами хряща, но и заходят в межкольцевые промежутки хрящевой части трахеи и в большей степени главных бронхов. Таким образом, в трахее пучки гладких мышц с поперечным и косым расположением находятся только в мембранозной части, т. е. мышечный слой как таковой отсутствует. В главных бронхах редкие группы гладких мышц имеются по всей окружности.
С уменьшением диаметра бронхов мышечный слой становится сильнее развитым, а волокна его идут в несколько косом направлении. Сокращение мышц вызывает не только с у -жение просвета бронхов, но и некоторое укорочение их, благодаря чему бронхи участвуют в выдохе за счет сокращения емкости дыхательных путей. Сокращение мышц позволяет сузить просвет бронхов на 1/4. При вдохе бронх удлиняется и расширяется. Мышцы достигают респираторных бронхиол 2-го порядка.
Кнутри от мышечного слоя находится подслизистый слой, состоящий из рыхлой соединительной ткани. В нем располагаются сосудистые и нервные образования, подслизистая лимфатическая сеть, лимфоидная ткань и значительная часть бронхиальных желез, которые относятся к трубчато-ацинозному типу со смешанной слизисто-серозной секрецией. Они состоят из концевых отделов и выводных протоков, которые открываются колбовидными расширениями на поверхности слизистой оболочки. Сравнительно большая длина протоков способствует длительному течению бронхитов при воспалительных процессах в железах. Атрофия желез может привести к высыханию слизистой оболочки и воспалительным изменениям.
Наибольшее число крупных желез имеется над бифуркацией трахеи и в области деления главных бронхов на долевые бронхи. У здорового человека в сутки выделяется до 100 мл секрета. На 95% он состоит из воды, а на 5% приходится равное количество белков, солей, липидов и неорганических веществ. В секрете преобладают муцины (высокомолекулярные гликопротеины). К настоящему времени насчитывается 14 видов гликопротеинов, 8 из которых содержатся в респираторной системе.
Слизистая оболочка бронхов
Слизистая оболочка состоит из покровного эпителия, базальной мембраны, собственной пластинки слизистой оболочки и мышечной пластинки слизистой оболочки.
Бронхиальный эпителий содержит высокие и низкие базальные клетки, каждая из которых прикреплена к базальной мембране. Толщина базальной мембраны колеблется от 3,7 до 10,6 мкм. Эпителий трахеи и крупных бронхов многорядный, цилиндрический, мерцательный. Толщина эпителия на уровне сегментарных бронхов составляет от 37 до 47 мкм. В его составе различают 4 основных типа клбток: реснитчатые, бокаловидные, промежуточные и базальные. Кроме того, встречаются серозные, щеточные, клетки Клара и Кульчицкого.
Реснитчатые клетки преобладают на свободной поверхности эпителиального пласта (Романова Л.К., 1984). Они имеют неправильную призматическую форму и овальное пузырьковидное ядро, расположенное в средней части клетки. Электроннооптическая плотность цитоплазмы невелика. Митохондрий немного, эндоплазматический гранулярный ретикулум развит слабо. Каждая клетка несет на своей поверхности короткие микроворсинки и около 200 мерцательных ресничек толщиной 0,3 мкм и длиной около 6 мкм. У человека плотность расположения ресничек составляет 6 мкм2.
Между соседними клетками образуются пространства; между собой клетки соединяются с помощью пальцеобразных выростов цитоплазмы и десмосом.
Популяция реснитчатых клеток по степени дифференцировки их апикальной поверхности подразделяется на следующие группы:
- Клетки, находящиеся в фазе формирования базальных телец и аксонем. Реснички в это время на апикальной поверхности отсутствуют. В этот период происходит накопление центриолей, которые перемещаются к апикальной поверхности клеток, и формирование базальных телец, из которых начинают образовываться аксонемы ресничек.
- Клетки, находящиеся в фазе умеренно выраженного цилиогенеза и роста ресничек. На апикальной поверхности таких клеток появляется небольшое количество ресничек, длина которых составляет 1/2-2/3 от длины ресничек дифференцированных клеток. В этой фазе на апикальной поверхности преобладают микроворсинки.
- Клетки, находящиеся в фазе активного цилиогенеза и роста ресничек. Апикальная по-верхность таких клеток уже почти целиком покрыта ресничками, размеры которых соответствуют размерам ресничек клеток, находящихся в предшествующей фазе цилиогенеза.
- Клетки, находящиеся в фазе завершенного цилиогенеза и роста ресничек. Апикальная поверхность таких клеток целиком покрыта густо расположенными длинными ресничками. На электронограммах видно, что реснички рядом расположенных клеток ориентированы в одном направлении и изогнуты. Это является выражением мукоцилиарного транспорта.
Все эти группы клеток хорошо различимы на фотографиях, полученных с помощью световой электронной микроскопии (СЭМ).
Реснички прикреплены к базальным тельцам, находящимся в апикальной части клетки. Аксонема реснички образована микротрубочками, из которых 9 пар (дуплеты) расположены по периферии, а 2 единичных (синглеты) — в центре. Дуплеты и синглеты соединены некси-новыми фибриллами. На каждом из дуплетов с одной стороны имеются 2 короткие «ручки», в которых содержится АТФ-аза, участвующая в освобождении энергии АТФ. Благодаря такой структуре реснички ритмично колеблются с частотой 16-17 в направлении носоглотки.
Они перемещают слизистую пленку, покрывающую эпителий, со скоростью около 6 мм/мин, обеспечивая тем самым непрерывную дренажную функцию бронха.
Реснитчатые эпителиоциты, по мнению большинства исследователей, находятся на стадии конечной дифференцировки и не способны к делению митозом. Согласно современной концепции, базальные клетки являются предшественниками промежуточных клеток, которые могут дифференцироваться в реснитчатые клетки.
Бокаловидные клетки, как и реснитчатые, достигают свободной поверхности эпителиального пласта. В мембранозной части трахеи и крупных бронхов на долю реснитчатых клеток приходится до 70-80%, а на долю бокаловидных — не более 20-30%. В тех местах, где по периметру трахеи и бронхов имеются хрящевые полукольца, обнаруживаются зоны с разным соотношением реснитчатых и бокаловидных клеток:
- с преобладанием реснитчатых клеток;
- с почти равным соотношением реснитчатых и секреторных клеток;
- с преобладанием секреторных клеток;
- с полным или почти полным отсутствием реснитчатых клеток («безреснитчатые»).
Бокаловидные клетки являются одноклеточными железами мерокринового типа, выделяющими слизистый секрет. Форма клетки и расположение ядра зависят от фазы секреции и заполнения надъядерной части гранулами слизи, которые сливаются в более крупные гранулы и характеризуются малой электронной плотностью. Бокаловидные клетки имеют удлиненную форму, которая во время накопления секрета принимает вид бокала с основанием, расположенным на базальной мембране и интимно связанным с ней. Широкий конец клетки куполообразно выступает на свободной поверхности и снабжен микроворсинками. Цитоплазма электронноплотная, ядро округлое, эндоплазматическая сеть шероховатого типа, хорошо развита.
Бокаловидные клетки распределены неравномерно. При сканирующей электронной микроскопии было выявлено, что различные зоны эпителиального пласта содержат неоднородные участки, состоящие либо только из реснитчатых эпителиоцитов, либо только из секреторных клеток. Однако сплошные скопления бокаловидных клеток сравнительно немногочисленны. По периметру на срезе сегментарного бронха здорового человека имеются участки, где соотношение реснитчатых эпителиоцитов и бокаловидных клеток составляет 4:1-7:1, а в других областях это соотношение равно 1:1.
Число бокаловидных клеток уменьшается в бронхах дистально. В бронхиолах бокаловидные клетки замещаются клетками Клара, участвующими в выработке серозных компонентов слизи и альвеолярной гипофазы.
В мелких бронхах и бронхиолах бокаловидные клетки в норме отсутствуют, но могут появляться при патологии.
В 1986 г. чешские ученые изучали реакцию эпителия воздухоносных путей кроликов на пероральное введение различных муколитических веществ. Оказалось, что клетками-мишенями действия муколитиков служат бокаловидные клетки. После выведения слизи бокаловидные клетки, как правило, дегенерируют и постепенно удаляются из эпителия. Степень повреждения бокаловидных клеток зависит от введенного вещества: наибольший раздражающий эффект дает ласольван. После введения бронхолизина и бромгексина происходит массивная дифференцировка новых бокаловидных клеток в эпителии воздухоносных путей, следствием чего является гиперплазия бокаловидных клеток.
Базальные и промежуточные клетки расположены в глубине эпителиального пласта и не достигают свободной поверхности. Это наименее дифференцированные клеточные формы, за счет которых в основном осуществляется физиологическая регенерация. Форма промежуточных клеток удлиненная, базальных — неправильно-кубическая. У тех и других — округлое, богатое ДНК ядро и небольшое количество цитоплазмы, имеющей большую плотность в базальных клетках.
Базальные клетки способны давать начало как реснитчатым, так и бокаловидным клеткам.
Секреторные и реснитчатые клетки объединяются под названием «мукоцилиарный аппарат».
Процесс передвижения слизи в воздухоносных путях легких называется мукоцилиарным клиренсом. Функциональная эффективность МЦК зависит от частоты и синхронности движения ресничек мерцательного эпителия, а также, что очень важно, от характеристики и реологических свойств слизи, т. е. от нормальной секреторной способности бокаловидных клеток.
Серозные клетки немногочисленны, достигают свободной поверхности эпителия и отличаются мелкими электронноплотными гранулами белкового секрета. Цитоплазма также электронноплотная. Хорошо развиты митохондрии и шероховатый ретикулум. Ядро округлое, обычно находится в средней части клетки.
Секреторные клетки, или клетки Клара, наиболее многочисленны в мелких бронхах и бронхиолах. Они, как и серозные, содержат мелкие электронноплотные гранулы, но отличаются малой электронной плотностью цитоплазмы и преобладанием гладкого, эндоплаз-матического ретикулума. Округлое ядро находится в средней части клетки. Клетки Клара участвуют в образовании фосфолипидов и, возможно, в выработке сурфактанта. В условиях повышенного раздражения они, по-видимому, могут превращаться в бокаловидные клетки.
Щеточные клетки несут на свободной поверхности микроворсинки, но лишены ресничек. Цитоплазма их малой электронной плотности, ядро овальное, пузырьковидное. В руководстве Хэма А. и Кормака Д. (1982) они рассматриваются как бокаловидные клетки, выделившие свой секрет. Им приписывается множество функций: абсорбционная, сократительная, секреторная, хеморецепторная. Однако в воздухоносных путях человека они практически не исследованы.
Клетки Кульчицкого встречаются на всем протяжении бронхиального дерева в основании эпителиального пласта, отличаясь от базальных малой электронной плотностью цитоплазмы и наличием мелких гранул, которые выявляются под электронным микроскопом и под световым при импрегнации серебром. Их относят к нейросекреторным клеткам APUD — системы.
Под эпителием находится базальная мембрана, которая состоит из коллагеновых и неколлагеновых гликопротеидов; она обеспечивает поддержку и прикрепление эпителия, участвует в метаболизме и иммунологических реакциях. Состояние базальной мембраны и подлежащей соединительной ткани обусловливает структуру и функцию эпителия. Собственной пластинкой называют слой рыхлой соединительной ткани между базальной мембраной и мышечным слоем. В ней находятся фибробласты, коллагеновые и эластические волокна. В собственной пластинке имеются кровеносные и лимфатические сосуды. Капилляры достигают базальной мембраны, но не проникают в нее.
В слизистой оболочке трахеи и бронхов, преимущественно в собственной пластинке и возле желез, в подслизистой постоянно присутствуют свободные клетки, которые могут проникать через эпителий в просвет. Среди них преобладают лимфоциты, реже встречаются плазматические клетки, гистиоциты, тучные клетки (лаброциты), нейтрофильные и эозинофильные лейкоциты. Постоянное нахождение лимфоидных клеток в слизистой оболочке бронхов обозначается специальным термином «бронхоассоциированная лимфоидная ткань» (БАЛТ) и рассматривается в качестве иммунологической защитной реакции на антигены, проникающие с воздухом в дыхательные пути.
[11], [12], [13], [14], [15], [16], [17]
- Daremberg, «Histoire des sciences médicales» (П., 1966).
- Мустафин Р. И., Протасова А. А., Буховец А. В., Семина И.И. Исследование интерполимерных сочетаний на основе (мет)акрилатов в качестве перспективных носителей в поликомплексных системах для гастроретентивной доставки. Фармация. 2014; 5: 3–5.
- М.П. Киселева, З.С. Шпрах, Л.М. Борисова и др. Доклиническое изучение противоопухолевой активности производного N-гликозида индолокарбазола ЛХС-1208. Сообщение II // Российский биотерапевтический журнал. 2015. № 3. С. 41-47.
- https://meduniver.com/Medical/pulmonologia/217.html.
- https://meduniver.com/Medical/pulmonologia/123.html.
- https://meduniver.com/Medical/pulmonologia/100.html.
- https://ilive.com.ua/health/bronhi_110313i16006.html.
- ОФС.1.2.1.1.0003.15 Спектрофотометрия в ультрафиолетовой и видимой областях // Государственная фармакопея, XIII изд.









